
Rod Sclerocactus Britton et Rose (trochu i nak)
Zahrnout do: mrazuvzdorné kaktusy
Kaktusy 2000, č. 2
Moderná taxonómia sa zaoberá vedeckým triedením organizmov podľa fylogenetických vzťahov. Základnou jednotkou je druh (species), čo je súhrn jedincov jednotného pôvodu, so spoločnými charakteristickými znakmi a vlastnosťami, ktoré v aktuálnom čase a priestore prenášajú na svoje potomstvo a vyskytujú sa v určitej geografickej oblasti a v určitých ekologických podmienkach. V praxi sa druh ďalej delí na poddruh (subspecies), odrodu (varietas). Druhy sa zoskupujú do rodov (genus), čeľadí (familia) atď.

S. parviflorus SB 474 – jeden z najkrajších zástupcov rodu
Delenie rastlín do systému, teda systematických jednotiek, má svoje zákonitosti, ide o snahu zaradiť ich do fylogenetickej vývojovej rady podľa určitých vývojových znakov. Výsledky rôznych vedeckých oborov pomohli upresniť fylogenetický systém a boli presne určené znaky pre čeľaď, o niečo menej pre rod a najmenej pre druh či varietu. Deliace znaky pre rod sú opakujúcimi znakmi u rôznych druhov hoci i na nesúvislom priestore, naopak deliace znaky pre druh sú špecifické znaky, ktorými sa súbor jedincov odlišuje od iného súboru na ohraničenom priestore. Pokiaľ dôjde k odlišnostiam v rámci druhu, pri opakovaní sa odlišných znakov druhu sa vytvára predpoklad pre poddruh či varietu. Forma má odlišné znaky od ostatných znakov druhu, či variety, ale znaky nemusia byť dedičné. Formy na stanovišti majú svoje odlišné znaky dané podmienkami v prírode, kde sa istá populácia nachádza. Druhy, u ktorých dochádza k hybridizácii v prírode a k prechodným medzidruhovým formám, sú druhmi vo vývoji s neukončeným speciačným procesom. Rôznorodosť prírodných podmienok, v ktorých sa druh nachádza, vedie k väčšej či menšej variabilite a je ťažké určiť kvantitatívny interval deliacich znakov druhu. Žiada si to dôkladný poľný výskum a porovnávací výskum v rovnakých podmienkach.

S. polyancistrus (San Bernadino Co., CA)
Tých pár viet som úmyselne uviedol preto, že práve nie veľký rod Sclerocactus zápasí od vzniku svojho prvého zástupcu až dodnes s problémami ako určiť deliace znaky jednotlivých druhov v prostredí nesmierne rôznorodom, často s izolovanými populáciami, ktoré z hľadiska prírodných prekážok sa nemôžu rozširovať, alebo práve ich rozširovanie bolo ovplyvnené iba istými smermi. To, čo vskutku o tom rozhodovalo, sú formácie Skalnatých Hôr a toky, ktoré smerujú v smere sever–juh. Nemenej dôležité sú mikroklimatické podmienky jednotlivých kaňonov, ktoré ešte viac upresňujú tie podmienky, ktoré mali rozhodujúci vplyv na ďalšie delenie. Nemyslím si, že najväčším problémom je problém okruhov S. whipplei–S. parviflorus, ale vnútorný problém všetkých druhov, ktorý sa prejavuje veľkou variabilitou, kde je veľmi ťažké určiť priority taxonomických jednotiek.

S. parviflorus HK 1083 (terrae-canyonae)
Ak by sme sa nezáväzne pokúsili určiť deliace znaky rodu Sclerocactus, tak by sme uviedli asi nasledovne: „Stonka jednotlivá alebo odnožujúca, rebrá rozpadnuté v hrbolce, ktoré sú v hornej časti opatrené brázdou, na konci ktorej v blízkosti areoly je reprodukčný orgán a žľaza roniaca nektár, v tomto mieste sa môže tvoriť jeden kvet, alebo i jedna odnož, areoly sú kruhové až elipsovité, holé až v mladosti mierne plstnaté, okrajové tŕne sú lúčovité, stredové odstávajúce, rovné až ohnuté, často hákovité, nepočetné, kvet je holý, lievikovitý, okvetné lístky sú špirálovito usporiadané, plod je holý, na povrchu často so zaschnutými najnižšími okvetnými lístkami, ktoré sa javia ako šupinky, po zrelosti zasýcha, semená veľké, čierne, s tvrdou testou, s bradavičnatým povrchom. Areál výskytu je polopúšť Juhozápadu USA.“

S. parviflorus SB 734 – kvet žltý až bledo ružový
Pokiaľ hodnotíme taxonomickú nomenklatúru zástupcov rodu z pohľadu dneška, tak s úžasom zisťujeme, že mnohé znaky, ktoré boli uvedené v popisoch, sú nedostatočne definované, ba dokonca sú mylne uvedené. Pravdepodobne je to dôsledok nedostatočných znalostí o celej populácii rastlín toho istého druhu v čase popisu. Ak to hodnotíme z pohľadu doby, v ktorej ten či onen taxon bol popísaný, tak sa ani tomu nemôžeme diviť, pretože zástupcovia rodu sa nachádzali v oblastiach, kde poľný výskum bol životu nebezpečný a tak pre popis bola použitá často iba jediná rastlina náhodne objavená. V sedemdesiatych rokoch tohoto storočia bol vykonaný poľný výskum zameraný hlavne na rod Sclerocactus profesionálmi okolo L. Bensona, ale nedošlo k objasneniu problémov v rode. Naopak bolo popísaných viac taxonov, u ktorých neboli dostatočne definované hodnoty taxonomickej kategórie, čím došlo skôr k zahmleniu ako k objasneniu problémov v rode. Táto doba pripomínala skôr preteky ako serióznu botanickú prácu. V poslednom období v teréne pracovalo viac nie botanikov, ktorí oveľa dôkladnejšie spoznali ekológiu druhov, no nazdávam sa, že výsledok ich práce sa dá označiť iba za snahu vniesť do problému jasno, s neurčitým výsledkom. Tým myslím hlavne na F. Hochstättera, asi toho času najviac poznajúceho prírodné stanovištia, G. Haslingera a A. Muehla. Nám nič nezostáva iba si počkať na serióznu revíziu rodu a pre istotu naďalej uvádzať pri názvoch i poľné čísla.

S. parviflorus SB 848 – patrí medzi „veľkokveté″
Už niekoľko rokov sledujem mne dostupné taxony v mojej zbierke od výsevu až po kvitnutie. Som si vedomý, že materiál nie je postačujúci, aby vystihoval celú škálu rodu, na druhej strane sú to zástupcovia, ktorí predsa len čo to napovedajú a poskytujú vedomosti o rôznych druhoch v rovnakých podmienkach. Niektoré taxony už niekoľko desaťročí kolujú v našich zbierkach aj s prípadnými chybami v názvoch, čo z hľadiska botanického považujem za takmer bezcenný materiál. Z hľadiska zberateľského sú to veľmi zaujímavé kaktusy a môžeme si overiť hlavne svoju pestovateľskú zdatnosť.

S. parviflorus GH 88/B02-02 pôvod Moab, Grand Co., UT
Rok 1999 z hľadiska pestovateľského som hodnotil ako divný a skôr neúspešný z dôvodu, že niektoré kaktusy, ktoré mi v minulosti kvitli bohato, teraz buď vôbec nekvitli, alebo kvitli s menším počtom kvetov. Výnimku tvorili zástupcovia rodu Sclerocactus, ktorí mi zakvitli bohato a poskytli mi vzácne poznatky. Zdokumentoval som hlavne veľkosti kvetov a u plodov som sa zameral hlavne na spôsob otvárania.
Pokúsim sa uviesť niektoré skutočnosti v poradí od konca. Semená vykazovali rôzne veľkosti a to podľa počtu semien v plode. Čím viac semien bolo v plode, tým boli menšie. I napriek tomu medzi tie s väčšími semenami patria S. wrightiae L. Benson, S. mesae-verdae L. Benson, S. polyancistrus Britton et Rose a S. nyensis Hochstätter. Tu by som chcel pripomenúť, že rastliny, ktoré pochádzajú z Utahu a majú tiež veľké semená v našich zbierkach kolovali ako S. wrightiae s ružovým kvetom. Pán F. Hochstätter ich zaradil ako S. parviflorus ssp. parviflorus f. macrospermus. Už pri menšej dávke fantázie by sme ich mohli považovať za istú prechodnú formu S. wrightiae.

S. whipplei SB 472 – typický zástupca druhu
Pokiaľ sa v popisoch tvar plodu uvádza väčšinou ako guľovitý, v skutočnosti sa jedná vo väčšine o tvar predĺžený, teda elipsoidný až tvar olivy. Je potrebné rozlišovať počas dozrievania plodu isté fázy, z ktorých je pre nás najdôležitejšia doba zrelosti a spôsob uvoľňovania semien z plodu. Plody sa v dobe zrelosti buď otvárajú pukaním, keď sa z plodu uvoľňujú semená, alebo plod sa neotvára od seba, iba v dôsledku mechanického poškodenia hlavne hmyzom, ktorý je lákaný sladkou šťavou oplodia. Plody po puknutí sa ľahko oddeľujú a odpadávajú, čím sa semená vysemeňujú. Plody, ktoré nepukajú, prechádzajú do štádia zasýchania, keď sa postupne zbavujú vody v oplodí, pričom sa oddeľujú rôzne. Niektoré ťažko, niektoré ľahko. Tie, ktoré po zasušení zostanú tvrdé, sa oddeľujú ľahko. Tie, čo po zasušení zostávajú mäkké z dôvodu, že oplodie zostane blanovité, sa oddeľujú ťažko. Tvrdé plody mali S. parviflorus Clover et Jotter (SB 593, SB 734, SB 474, SB 132, SB 592), ostatné mali viac menej mäkké plody. Úplne jednoznačne sa chovali plody S. glaucus L. Benson. Tie po dozretí odparili vodu z oplodia a tak zostali iba blanovité, jemne ryhované, priesvitné oplodia s obrysmi semien. Tieto plody sa sami od seba neotvárali, v tomto stave vydržali i do jari. Mali zvláštny denný „režim“. Ráno boli mäkké a pružné po tom, čo bola vyššia vlhkosť vzduchu, cez deň sa vysušili a boli tvrdé a lámavé. Po mechanickom poškodení sa semená uvoľňovali. Toto cyklovanie má pravdepodobne vplyv i na klíčivosť semien. Najväčšie rozdiely voči popisom sa našli pri údajoch o otváraní plodov. Pokiaľ sa iba u S. polyancistrus uvádza, že sa otvára horizontálne, teda vertikálnou puklinou, tak u ostatných sa uvádza, že sa jedná o nepravidelné pukanie. V skutočnosti dochádzalo k pukaniu od dolnej časti pravidelnou vertikálnou puklinou u S. polyancistrus, S. nyensis, S. pubispinus, S. spinosior, S. schleseri Heil et Welsh. U S. mesae-verdae dochádzalo k nepravidelnému horizontálnemu pukaniu v dolnej, alebo hornej časti plodu. U ostatných plodov nedochádzalo k pukaniu plodov, i keď sa semená sem-tam uvoľňovali po poškodení plodu mravcami a osami. Bolo evidentné, že mravce rýchlo nachádzali dozrievajúce plody u rastlín umiestnených vonku volne a narušovali plod od spodnej časti menšími dierkami, cez ktoré vynášali semená. Často načínali už aj nezrelé plody. Rastliny tých istých zástupcov, ktoré boli v skleníku, nemali porušené plody. Osi vyhrýzali plody od hora. Tieto plody sami od seba nepukali!

S. polyancistrus forma „albino″
Ďalšie údaje, ktoré považujem v popisoch za nepresné, sú údaje o kvetoch. Pre mňa je nepochopiteľné, keď profesionálni botanici uvádzali, že kvety kaktusov majú vonkajšie kališné lístky – sepálie – a korunné lupienky – petálie. Ani v ďalších upresneniach tieto údaje neboli uvedené na správnu mieru. V skutočnosti kvet odzrkadľuje taxonomické hodnoty všetkých kategórií. Prekonáva isté etapy fylogenetického vývoja od nasadenia puku až po rozkvitnutie. Dôkazom toho sú i rôzne tvary, veľkosti a okraje okvetných lístkov na tom istom kvete, ale aj zväčšujúca sa veľkosť počas kvitnutia. Kvety sú petaloidné, bez rozlíšenia okvetných lístkov, ich usporiadanie je v špirále v zostupnom poradí, so spodným semenníkom, ktorý je holý, taktiež so špirálovitým usporiadaním tyčiniek. Tvar sa v popisoch uvádza okrem lievikovitého aj rúrkovitý (S. spinosior Woodroff et Benson 1976, S. schleseri Hochstätter 1992, S. nyensis Hochstätter 1992, S. wetlandicus ssp. ilseae Hochstätter 1993, S. whipplei ssp. busekii Hochstätter 1995), lievikovitý až rúrkovitý (S. parviflorus Clover et Jotter 1941), ba dokonca zvonovitý (S. parviflorus ssp. havasupaiensis Hochstätter 1995). V skutočnosti sa u všetkých druhov jedná o kvet lievikovitý, čo považujem za výrazný znak rodu. Medzi rozlišovacie znaky druhov považujem aj veľkosť kvetov. Najväčší kvet má S. polyancistrus, najmenší kvet S. wetlandicus ssp. ilseae, čo môže byť tiež znakom fylogenetického vývoja, ktorý vyjadruje stupeň, alebo vek vzniku druhu. Pochopiteľne ten najväčší má najprimitívnejší druh v rebríčku rodu. Farba kvetov je dedičná, a pre ten ktorý druh je charakteristická. Iba výnimočne dochádza k farbe s prechodným odtieňom. Všetky ostatné znaky kvetu nie sú dostatočne v popisoch dokumentované. Skutočne by bolo potrebné podchytiť údaje čo najväčšieho počtu rastlín týkajúcich sa farby a počtu ramien blizny, farby čnelky, nitky atď. Uvediem príklad. U väčšiny rastlín S. wrightiae, ktoré som videl kvitnúť, boli nitky hnedo-fialové, iba u jednej boli žlté, u mňa odhadujem na pomer 20:1, i napriek tomu sa uvádzajú ako žlté.

S. nyensis fh 105 – redšie a dlhšie tŕne
Absolútne chýbajú údaje o vzájomnom usporiadaní jednotlivých častí kvetu. Všimol som si ešte jednu skutočnosť, a to, že skupina rastlín, ktorá pochádzala z približne rovnakého prostredia, kvitla v jednom termíne. Ostatné skupiny kvitli v posunutých termínoch.

S. wrightiae (Wayne Co., UT)
Pokiaľ budeme hľadať súvislosti a zákonitosti v množstve a usporiadaní tŕňov, máme dve možnosti. Tá prvá je spochybniť správnosť zaradenia S. mesae-verdae do rodu Sclerocactus. Tá druhá je brať veľmi volne všetky údaje o tŕňoch a hľadať postupnosť fylogenetického vývoja u všetkých druhov. Tak či onak môžeme vysloviť isté zásady, ktoré platia s istými výnimkami pre všetky druhy. Okrajové tŕne sú lúčovito usporiadané okolo areoly, ich počet je okolo 10 a výnimočne u niektorých jedincov dosahuje až 15 – u S. polyancistrus a S. nyensis. Ďalším pravidlom je, že sa nachádza minimálne jeden plochý tŕň, rôzne dlhý, ktorý smeruje smerom hore, niekedy do boku, na povrchu je ryhovaný, a v popisoch sa uvádza ako „papierovitý“. Ak berieme do úvahy celý rod, tak výnimočne môže chýbať. Pôvodne som s týmto označením nesúhlasil, lebo moja predstava o takomto tŕni bola spojená s T. papyracantha. Po tom, čo som mal rastliny umiestnené vonku, kde na ne pršalo, som zistil, že v dobe väčšej vlhkosti tieto tŕne boli mäkké a flexibilné, iba v suchom stave boli tvrdé a pichľavé. Ich počet je od 0 (S. mesae-verdae) až 3 (S. polyancistrus, S. nyensis, S. schleseri). Ďalším pravidlom je, že sa jeden stredový hákovitý tŕň nachádza v strede areoly, kolmý na ňu. Aj tento výnimočne u niektorých jedincov môže chýbať. Ďalšie stredové tŕne sú viac menej znakom druhovým. Jedná sa o dva horné a dva dolné rovné, alebo hákovité odstávajúce do bokov. Ich počet a dĺžka sa odlišuje už v jednom výseve toho istého taxonu. V našich zbierkach dochádzalo k omylom v súvislosti s počtom stredových tŕňov. Poznal som jedného kaktusára, ktorý vyraďoval všetky rastliny S. glaucus, ktoré mali hákovité stredové tŕne, pretože to považoval za znaky hybridizácie. V tomto prípade sa jedná o znak formy zo stanovišťa. Skôr jedinci bez stredových tŕňov sú výnimkou. Publikoval sa obrázok, na ktorom bol údajne hybrid S. whipplei a S. mesae-verdae, v skutočnosti tam bola rastlina, ktorá zodpovedala SB 1011, rastline z Nového Mexika, u ktorých výskyt bez stredových tŕňov bol v pomere k ostatným cca 1:1. Ja len neviem, čo potom s tým malým fialovým kvetom, keď rastlina zakvitla. Už dlhodobo používam také nepísané pravidlo týkajúce sa rozlíšenia S. whipplei od S. parviflorus. Ide o zásadu, keď u prv spomínaného, stredový plochý tŕň je najväčší zo všetkých tŕňov. U S. parviflorus ten istý tŕň je kratší ako najdlhší stredový. Ďalším rozlišovacím znakom sú znaky fylogenetického vývoja. Je známe, že aj kaktusy od vyklíčenia až po dospelosť prekonávajú svoj fylogenetický vývoj. U S. whipplei sa v mladosti vyvíjajú iba okrajové tŕne, až v neskoršom veku sa vyvíjajú stredové. U S. parviflorus sa stredové tŕne vyvíjajú už od mladého veku. Tak isto je možné konštatovať, že páperovité tŕne v mladom veku S. pubispinus, S. spinosior, S. schleseri súvisia s úzkou príbuznosťou, tie isté u S. glaucus, kde vzhľadom na odlišné geografické rozšírenie sa pravdepodobne jedná o vznik taxonov v približne rovnakom období vývoja. Údaje o počte, dĺžke, farbe a usporiadaní tŕňov majú menšiu taxonomickú hodnotu ako údaje o kvetoch. To si môžeme overiť i sami, pretože na zmenené podmienky rastliny reagujú hlavne kvantitatívnymi zmenami na tŕňoch.

S. wetlandicus GH 88/B03-03

S. glaucus (Mesa Co., CO)
Mojou snahou nebolo komentovať delenie rodu dnes, ale bolo uviesť časť z rozsiahleho materiálu, ktorý mám k dispozícii, a upozorniť na tie nezrovnalosti, ktoré považujem za dôležité a možná i zaujímavé. Je pochopiteľné, že to, k čomu som dospel, vzniklo na základe zistených skutočností v mojom prostredí tak, ako sa to javilo mne bezprostredne po tom, čo som to zdokumentoval. Poznatky o čo najširšej populácii môžu byť rozdielne, pretože by sa týkali rozdielnych podmienok, no spôsob otvárania plodov by sa nemal meniť. Som presvedčený, že ešte zostáva veľa miest v prírode USA, ktoré prinesú nejedno prekvapenie a pomôžu objasniť tie problémy, ktoré dnes za problémy považujeme. Taktiež som presvedčený, že nevyjasnené skutočnosti týkajúce sa rodu Sclerocactus budú objasnené iba s prihliadnutím na fylogenetický vývoj rodu, s istou dávkou veľkorysosti tak, ako si to tieto skutočné skvosty zaslúžia.
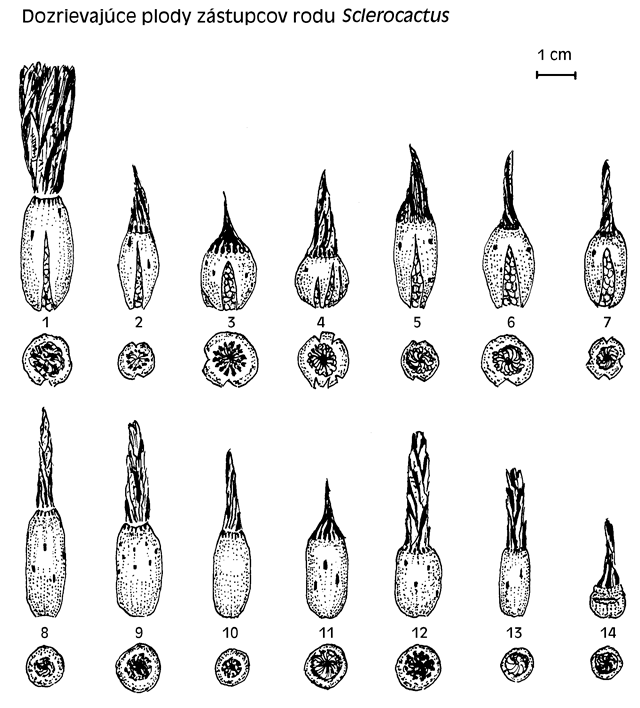
Dozreievajúce plody zástupcov rodu Sclerocactus
1. S. polyancistrus (San Bernadino Co., CA)
2. S. nyensis fh 105
3. S. pubispinus SB 1085
4. S. spinosior JB 22
5. S. spinosior SB 740
6. S. spinosior SB 1182
7. S. schleseri SB 1045
8. S. parviflorus SB 593
9. S. parviflorus SB 734
10. S. parviflorus GH 91/B08-06
11. S. whipplei GH 91/B08-5
12. S. parviflorus SB 474
13. S. whipplei v. pygmaeus
14. S. mesae-verdae
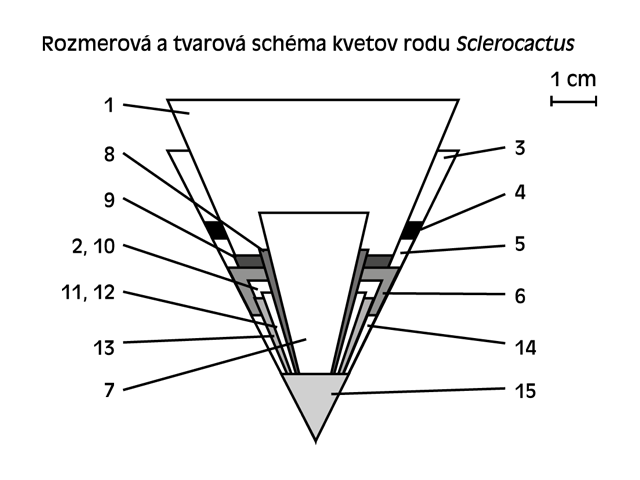
1. S. polyancistrus (San Bernadino Co., CA)
2. S. nyensis fh 105
3. S. parviflorus SB 848
4. S. parviflorus (terrae-canyonae) SB 592
5. S. parviflorus SB 734
6. S. af. whipplei-parviflorus SB 1011
7. S. whipplei SB 736
8. S. wrightiae fh 69.7
9. S. glaucus GH 91/B02-06
10. S. mesae-verdae (Montezuma Co., CO)
11. S. schleseri SB 1085
12. S. spinosior GH 91/B06-07
13. S. pubispinus SB 1085
14. S. wetlandicus GH 88/B03-03
15. S. wetlandicus v. ilseae (UT)
Moderná taxonómia sa zaoberá vedeckým triedením organizmov podľa fylogenetických vzťahov. Základnou jednotkou je druh (species), čo je súhrn jedincov jednotného pôvodu, so spoločnými charakteristickými znakmi a vlastnosťami, ktoré v aktuálnom čase a priestore prenášajú na svoje potomstvo a vyskytujú sa v určitej geografickej oblasti a v určitých ekologických podmienkach. V praxi sa druh ďalej delí na poddruh (subspecies), odrodu (varietas). Druhy sa zoskupujú do rodov (genus), čeľadí (familia) atď.
S. parviflorus SB 474 – jeden z najkrajších zástupcov rodu
Delenie rastlín do systému, teda systematických jednotiek, má svoje zákonitosti, ide o snahu zaradiť ich do fylogenetickej vývojovej rady podľa určitých vývojových znakov. Výsledky rôznych vedeckých oborov pomohli upresniť fylogenetický systém a boli presne určené znaky pre čeľaď, o niečo menej pre rod a najmenej pre druh či varietu. Deliace znaky pre rod sú opakujúcimi znakmi u rôznych druhov hoci i na nesúvislom priestore, naopak deliace znaky pre druh sú špecifické znaky, ktorými sa súbor jedincov odlišuje od iného súboru na ohraničenom priestore. Pokiaľ dôjde k odlišnostiam v rámci druhu, pri opakovaní sa odlišných znakov druhu sa vytvára predpoklad pre poddruh či varietu. Forma má odlišné znaky od ostatných znakov druhu, či variety, ale znaky nemusia byť dedičné. Formy na stanovišti majú svoje odlišné znaky dané podmienkami v prírode, kde sa istá populácia nachádza. Druhy, u ktorých dochádza k hybridizácii v prírode a k prechodným medzidruhovým formám, sú druhmi vo vývoji s neukončeným speciačným procesom. Rôznorodosť prírodných podmienok, v ktorých sa druh nachádza, vedie k väčšej či menšej variabilite a je ťažké určiť kvantitatívny interval deliacich znakov druhu. Žiada si to dôkladný poľný výskum a porovnávací výskum v rovnakých podmienkach.
S. polyancistrus (San Bernadino Co., CA)
Tých pár viet som úmyselne uviedol preto, že práve nie veľký rod Sclerocactus zápasí od vzniku svojho prvého zástupcu až dodnes s problémami ako určiť deliace znaky jednotlivých druhov v prostredí nesmierne rôznorodom, často s izolovanými populáciami, ktoré z hľadiska prírodných prekážok sa nemôžu rozširovať, alebo práve ich rozširovanie bolo ovplyvnené iba istými smermi. To, čo vskutku o tom rozhodovalo, sú formácie Skalnatých Hôr a toky, ktoré smerujú v smere sever–juh. Nemenej dôležité sú mikroklimatické podmienky jednotlivých kaňonov, ktoré ešte viac upresňujú tie podmienky, ktoré mali rozhodujúci vplyv na ďalšie delenie. Nemyslím si, že najväčším problémom je problém okruhov S. whipplei–S. parviflorus, ale vnútorný problém všetkých druhov, ktorý sa prejavuje veľkou variabilitou, kde je veľmi ťažké určiť priority taxonomických jednotiek.
S. parviflorus HK 1083 (terrae-canyonae)
Ak by sme sa nezáväzne pokúsili určiť deliace znaky rodu Sclerocactus, tak by sme uviedli asi nasledovne: „Stonka jednotlivá alebo odnožujúca, rebrá rozpadnuté v hrbolce, ktoré sú v hornej časti opatrené brázdou, na konci ktorej v blízkosti areoly je reprodukčný orgán a žľaza roniaca nektár, v tomto mieste sa môže tvoriť jeden kvet, alebo i jedna odnož, areoly sú kruhové až elipsovité, holé až v mladosti mierne plstnaté, okrajové tŕne sú lúčovité, stredové odstávajúce, rovné až ohnuté, často hákovité, nepočetné, kvet je holý, lievikovitý, okvetné lístky sú špirálovito usporiadané, plod je holý, na povrchu často so zaschnutými najnižšími okvetnými lístkami, ktoré sa javia ako šupinky, po zrelosti zasýcha, semená veľké, čierne, s tvrdou testou, s bradavičnatým povrchom. Areál výskytu je polopúšť Juhozápadu USA.“
S. parviflorus SB 734 – kvet žltý až bledo ružový
Pokiaľ hodnotíme taxonomickú nomenklatúru zástupcov rodu z pohľadu dneška, tak s úžasom zisťujeme, že mnohé znaky, ktoré boli uvedené v popisoch, sú nedostatočne definované, ba dokonca sú mylne uvedené. Pravdepodobne je to dôsledok nedostatočných znalostí o celej populácii rastlín toho istého druhu v čase popisu. Ak to hodnotíme z pohľadu doby, v ktorej ten či onen taxon bol popísaný, tak sa ani tomu nemôžeme diviť, pretože zástupcovia rodu sa nachádzali v oblastiach, kde poľný výskum bol životu nebezpečný a tak pre popis bola použitá často iba jediná rastlina náhodne objavená. V sedemdesiatych rokoch tohoto storočia bol vykonaný poľný výskum zameraný hlavne na rod Sclerocactus profesionálmi okolo L. Bensona, ale nedošlo k objasneniu problémov v rode. Naopak bolo popísaných viac taxonov, u ktorých neboli dostatočne definované hodnoty taxonomickej kategórie, čím došlo skôr k zahmleniu ako k objasneniu problémov v rode. Táto doba pripomínala skôr preteky ako serióznu botanickú prácu. V poslednom období v teréne pracovalo viac nie botanikov, ktorí oveľa dôkladnejšie spoznali ekológiu druhov, no nazdávam sa, že výsledok ich práce sa dá označiť iba za snahu vniesť do problému jasno, s neurčitým výsledkom. Tým myslím hlavne na F. Hochstättera, asi toho času najviac poznajúceho prírodné stanovištia, G. Haslingera a A. Muehla. Nám nič nezostáva iba si počkať na serióznu revíziu rodu a pre istotu naďalej uvádzať pri názvoch i poľné čísla.
S. parviflorus SB 848 – patrí medzi „veľkokveté″
Už niekoľko rokov sledujem mne dostupné taxony v mojej zbierke od výsevu až po kvitnutie. Som si vedomý, že materiál nie je postačujúci, aby vystihoval celú škálu rodu, na druhej strane sú to zástupcovia, ktorí predsa len čo to napovedajú a poskytujú vedomosti o rôznych druhoch v rovnakých podmienkach. Niektoré taxony už niekoľko desaťročí kolujú v našich zbierkach aj s prípadnými chybami v názvoch, čo z hľadiska botanického považujem za takmer bezcenný materiál. Z hľadiska zberateľského sú to veľmi zaujímavé kaktusy a môžeme si overiť hlavne svoju pestovateľskú zdatnosť.
S. parviflorus GH 88/B02-02 pôvod Moab, Grand Co., UT
Rok 1999 z hľadiska pestovateľského som hodnotil ako divný a skôr neúspešný z dôvodu, že niektoré kaktusy, ktoré mi v minulosti kvitli bohato, teraz buď vôbec nekvitli, alebo kvitli s menším počtom kvetov. Výnimku tvorili zástupcovia rodu Sclerocactus, ktorí mi zakvitli bohato a poskytli mi vzácne poznatky. Zdokumentoval som hlavne veľkosti kvetov a u plodov som sa zameral hlavne na spôsob otvárania.
Pokúsim sa uviesť niektoré skutočnosti v poradí od konca. Semená vykazovali rôzne veľkosti a to podľa počtu semien v plode. Čím viac semien bolo v plode, tým boli menšie. I napriek tomu medzi tie s väčšími semenami patria S. wrightiae L. Benson, S. mesae-verdae L. Benson, S. polyancistrus Britton et Rose a S. nyensis Hochstätter. Tu by som chcel pripomenúť, že rastliny, ktoré pochádzajú z Utahu a majú tiež veľké semená v našich zbierkach kolovali ako S. wrightiae s ružovým kvetom. Pán F. Hochstätter ich zaradil ako S. parviflorus ssp. parviflorus f. macrospermus. Už pri menšej dávke fantázie by sme ich mohli považovať za istú prechodnú formu S. wrightiae.
S. whipplei SB 472 – typický zástupca druhu
Pokiaľ sa v popisoch tvar plodu uvádza väčšinou ako guľovitý, v skutočnosti sa jedná vo väčšine o tvar predĺžený, teda elipsoidný až tvar olivy. Je potrebné rozlišovať počas dozrievania plodu isté fázy, z ktorých je pre nás najdôležitejšia doba zrelosti a spôsob uvoľňovania semien z plodu. Plody sa v dobe zrelosti buď otvárajú pukaním, keď sa z plodu uvoľňujú semená, alebo plod sa neotvára od seba, iba v dôsledku mechanického poškodenia hlavne hmyzom, ktorý je lákaný sladkou šťavou oplodia. Plody po puknutí sa ľahko oddeľujú a odpadávajú, čím sa semená vysemeňujú. Plody, ktoré nepukajú, prechádzajú do štádia zasýchania, keď sa postupne zbavujú vody v oplodí, pričom sa oddeľujú rôzne. Niektoré ťažko, niektoré ľahko. Tie, ktoré po zasušení zostanú tvrdé, sa oddeľujú ľahko. Tie, čo po zasušení zostávajú mäkké z dôvodu, že oplodie zostane blanovité, sa oddeľujú ťažko. Tvrdé plody mali S. parviflorus Clover et Jotter (SB 593, SB 734, SB 474, SB 132, SB 592), ostatné mali viac menej mäkké plody. Úplne jednoznačne sa chovali plody S. glaucus L. Benson. Tie po dozretí odparili vodu z oplodia a tak zostali iba blanovité, jemne ryhované, priesvitné oplodia s obrysmi semien. Tieto plody sa sami od seba neotvárali, v tomto stave vydržali i do jari. Mali zvláštny denný „režim“. Ráno boli mäkké a pružné po tom, čo bola vyššia vlhkosť vzduchu, cez deň sa vysušili a boli tvrdé a lámavé. Po mechanickom poškodení sa semená uvoľňovali. Toto cyklovanie má pravdepodobne vplyv i na klíčivosť semien. Najväčšie rozdiely voči popisom sa našli pri údajoch o otváraní plodov. Pokiaľ sa iba u S. polyancistrus uvádza, že sa otvára horizontálne, teda vertikálnou puklinou, tak u ostatných sa uvádza, že sa jedná o nepravidelné pukanie. V skutočnosti dochádzalo k pukaniu od dolnej časti pravidelnou vertikálnou puklinou u S. polyancistrus, S. nyensis, S. pubispinus, S. spinosior, S. schleseri Heil et Welsh. U S. mesae-verdae dochádzalo k nepravidelnému horizontálnemu pukaniu v dolnej, alebo hornej časti plodu. U ostatných plodov nedochádzalo k pukaniu plodov, i keď sa semená sem-tam uvoľňovali po poškodení plodu mravcami a osami. Bolo evidentné, že mravce rýchlo nachádzali dozrievajúce plody u rastlín umiestnených vonku volne a narušovali plod od spodnej časti menšími dierkami, cez ktoré vynášali semená. Často načínali už aj nezrelé plody. Rastliny tých istých zástupcov, ktoré boli v skleníku, nemali porušené plody. Osi vyhrýzali plody od hora. Tieto plody sami od seba nepukali!
S. polyancistrus forma „albino″
Ďalšie údaje, ktoré považujem v popisoch za nepresné, sú údaje o kvetoch. Pre mňa je nepochopiteľné, keď profesionálni botanici uvádzali, že kvety kaktusov majú vonkajšie kališné lístky – sepálie – a korunné lupienky – petálie. Ani v ďalších upresneniach tieto údaje neboli uvedené na správnu mieru. V skutočnosti kvet odzrkadľuje taxonomické hodnoty všetkých kategórií. Prekonáva isté etapy fylogenetického vývoja od nasadenia puku až po rozkvitnutie. Dôkazom toho sú i rôzne tvary, veľkosti a okraje okvetných lístkov na tom istom kvete, ale aj zväčšujúca sa veľkosť počas kvitnutia. Kvety sú petaloidné, bez rozlíšenia okvetných lístkov, ich usporiadanie je v špirále v zostupnom poradí, so spodným semenníkom, ktorý je holý, taktiež so špirálovitým usporiadaním tyčiniek. Tvar sa v popisoch uvádza okrem lievikovitého aj rúrkovitý (S. spinosior Woodroff et Benson 1976, S. schleseri Hochstätter 1992, S. nyensis Hochstätter 1992, S. wetlandicus ssp. ilseae Hochstätter 1993, S. whipplei ssp. busekii Hochstätter 1995), lievikovitý až rúrkovitý (S. parviflorus Clover et Jotter 1941), ba dokonca zvonovitý (S. parviflorus ssp. havasupaiensis Hochstätter 1995). V skutočnosti sa u všetkých druhov jedná o kvet lievikovitý, čo považujem za výrazný znak rodu. Medzi rozlišovacie znaky druhov považujem aj veľkosť kvetov. Najväčší kvet má S. polyancistrus, najmenší kvet S. wetlandicus ssp. ilseae, čo môže byť tiež znakom fylogenetického vývoja, ktorý vyjadruje stupeň, alebo vek vzniku druhu. Pochopiteľne ten najväčší má najprimitívnejší druh v rebríčku rodu. Farba kvetov je dedičná, a pre ten ktorý druh je charakteristická. Iba výnimočne dochádza k farbe s prechodným odtieňom. Všetky ostatné znaky kvetu nie sú dostatočne v popisoch dokumentované. Skutočne by bolo potrebné podchytiť údaje čo najväčšieho počtu rastlín týkajúcich sa farby a počtu ramien blizny, farby čnelky, nitky atď. Uvediem príklad. U väčšiny rastlín S. wrightiae, ktoré som videl kvitnúť, boli nitky hnedo-fialové, iba u jednej boli žlté, u mňa odhadujem na pomer 20:1, i napriek tomu sa uvádzajú ako žlté.
S. nyensis fh 105 – redšie a dlhšie tŕne
Absolútne chýbajú údaje o vzájomnom usporiadaní jednotlivých častí kvetu. Všimol som si ešte jednu skutočnosť, a to, že skupina rastlín, ktorá pochádzala z približne rovnakého prostredia, kvitla v jednom termíne. Ostatné skupiny kvitli v posunutých termínoch.
S. wrightiae (Wayne Co., UT)
Pokiaľ budeme hľadať súvislosti a zákonitosti v množstve a usporiadaní tŕňov, máme dve možnosti. Tá prvá je spochybniť správnosť zaradenia S. mesae-verdae do rodu Sclerocactus. Tá druhá je brať veľmi volne všetky údaje o tŕňoch a hľadať postupnosť fylogenetického vývoja u všetkých druhov. Tak či onak môžeme vysloviť isté zásady, ktoré platia s istými výnimkami pre všetky druhy. Okrajové tŕne sú lúčovito usporiadané okolo areoly, ich počet je okolo 10 a výnimočne u niektorých jedincov dosahuje až 15 – u S. polyancistrus a S. nyensis. Ďalším pravidlom je, že sa nachádza minimálne jeden plochý tŕň, rôzne dlhý, ktorý smeruje smerom hore, niekedy do boku, na povrchu je ryhovaný, a v popisoch sa uvádza ako „papierovitý“. Ak berieme do úvahy celý rod, tak výnimočne môže chýbať. Pôvodne som s týmto označením nesúhlasil, lebo moja predstava o takomto tŕni bola spojená s T. papyracantha. Po tom, čo som mal rastliny umiestnené vonku, kde na ne pršalo, som zistil, že v dobe väčšej vlhkosti tieto tŕne boli mäkké a flexibilné, iba v suchom stave boli tvrdé a pichľavé. Ich počet je od 0 (S. mesae-verdae) až 3 (S. polyancistrus, S. nyensis, S. schleseri). Ďalším pravidlom je, že sa jeden stredový hákovitý tŕň nachádza v strede areoly, kolmý na ňu. Aj tento výnimočne u niektorých jedincov môže chýbať. Ďalšie stredové tŕne sú viac menej znakom druhovým. Jedná sa o dva horné a dva dolné rovné, alebo hákovité odstávajúce do bokov. Ich počet a dĺžka sa odlišuje už v jednom výseve toho istého taxonu. V našich zbierkach dochádzalo k omylom v súvislosti s počtom stredových tŕňov. Poznal som jedného kaktusára, ktorý vyraďoval všetky rastliny S. glaucus, ktoré mali hákovité stredové tŕne, pretože to považoval za znaky hybridizácie. V tomto prípade sa jedná o znak formy zo stanovišťa. Skôr jedinci bez stredových tŕňov sú výnimkou. Publikoval sa obrázok, na ktorom bol údajne hybrid S. whipplei a S. mesae-verdae, v skutočnosti tam bola rastlina, ktorá zodpovedala SB 1011, rastline z Nového Mexika, u ktorých výskyt bez stredových tŕňov bol v pomere k ostatným cca 1:1. Ja len neviem, čo potom s tým malým fialovým kvetom, keď rastlina zakvitla. Už dlhodobo používam také nepísané pravidlo týkajúce sa rozlíšenia S. whipplei od S. parviflorus. Ide o zásadu, keď u prv spomínaného, stredový plochý tŕň je najväčší zo všetkých tŕňov. U S. parviflorus ten istý tŕň je kratší ako najdlhší stredový. Ďalším rozlišovacím znakom sú znaky fylogenetického vývoja. Je známe, že aj kaktusy od vyklíčenia až po dospelosť prekonávajú svoj fylogenetický vývoj. U S. whipplei sa v mladosti vyvíjajú iba okrajové tŕne, až v neskoršom veku sa vyvíjajú stredové. U S. parviflorus sa stredové tŕne vyvíjajú už od mladého veku. Tak isto je možné konštatovať, že páperovité tŕne v mladom veku S. pubispinus, S. spinosior, S. schleseri súvisia s úzkou príbuznosťou, tie isté u S. glaucus, kde vzhľadom na odlišné geografické rozšírenie sa pravdepodobne jedná o vznik taxonov v približne rovnakom období vývoja. Údaje o počte, dĺžke, farbe a usporiadaní tŕňov majú menšiu taxonomickú hodnotu ako údaje o kvetoch. To si môžeme overiť i sami, pretože na zmenené podmienky rastliny reagujú hlavne kvantitatívnymi zmenami na tŕňoch.
S. wetlandicus GH 88/B03-03
S. glaucus (Mesa Co., CO)
Mojou snahou nebolo komentovať delenie rodu dnes, ale bolo uviesť časť z rozsiahleho materiálu, ktorý mám k dispozícii, a upozorniť na tie nezrovnalosti, ktoré považujem za dôležité a možná i zaujímavé. Je pochopiteľné, že to, k čomu som dospel, vzniklo na základe zistených skutočností v mojom prostredí tak, ako sa to javilo mne bezprostredne po tom, čo som to zdokumentoval. Poznatky o čo najširšej populácii môžu byť rozdielne, pretože by sa týkali rozdielnych podmienok, no spôsob otvárania plodov by sa nemal meniť. Som presvedčený, že ešte zostáva veľa miest v prírode USA, ktoré prinesú nejedno prekvapenie a pomôžu objasniť tie problémy, ktoré dnes za problémy považujeme. Taktiež som presvedčený, že nevyjasnené skutočnosti týkajúce sa rodu Sclerocactus budú objasnené iba s prihliadnutím na fylogenetický vývoj rodu, s istou dávkou veľkorysosti tak, ako si to tieto skutočné skvosty zaslúžia.
Dozreievajúce plody zástupcov rodu Sclerocactus
1. S. polyancistrus (San Bernadino Co., CA)
2. S. nyensis fh 105
3. S. pubispinus SB 1085
4. S. spinosior JB 22
5. S. spinosior SB 740
6. S. spinosior SB 1182
7. S. schleseri SB 1045
8. S. parviflorus SB 593
9. S. parviflorus SB 734
10. S. parviflorus GH 91/B08-06
11. S. whipplei GH 91/B08-5
12. S. parviflorus SB 474
13. S. whipplei v. pygmaeus
14. S. mesae-verdae
1. S. polyancistrus (San Bernadino Co., CA)
2. S. nyensis fh 105
3. S. parviflorus SB 848
4. S. parviflorus (terrae-canyonae) SB 592
5. S. parviflorus SB 734
6. S. af. whipplei-parviflorus SB 1011
7. S. whipplei SB 736
8. S. wrightiae fh 69.7
9. S. glaucus GH 91/B02-06
10. S. mesae-verdae (Montezuma Co., CO)
11. S. schleseri SB 1085
12. S. spinosior GH 91/B06-07
13. S. pubispinus SB 1085
14. S. wetlandicus GH 88/B03-03
15. S. wetlandicus v. ilseae (UT)

